Chromozomy
Definice - co jsou chromozomy?
Genetická výbava buňky je uložena ve formě DNA (deoxyribonukleové kyseliny) a jejích bází (adenin, thymin, guanin a cytosin). Ve všech eukaryotických buňkách (zvířata, rostliny, houby) je tento přítomen v buněčném jádru ve formě chromozomů. Chromozom se skládá z jediné koherentní molekuly DNA, která je spojena s určitými proteiny.
Název chromozom je odvozen z řečtiny a lze jej zhruba přeložit jako „barevné tělo“. Tento název pochází ze skutečnosti, že velmi brzy v historii cytologie (1888) se vědcům podařilo barvit je pomocí speciálních základních barviv a identifikovat je ve světelném mikroskopu. Jsou však opravdu viditelné pouze v určitém bodě buněčného cyklu, mitózy (v zárodečných buňkách, meióza), kdy je chromozom obzvláště hustý (kondenzovaný).

Jak se vytvářejí chromozomy?
Pokud by byla propojena celá dvojitá šroubovice DNA buňky, tj. Přibližně 3,4 x 109 párů bází, vedlo by to k délce více než jednoho metru. Celková délka všech přidaných chromozomů je pouze asi 115 µm. Tento rozdíl v délce je vysvětlen velmi kompaktní strukturou chromozomů, ve kterých je DNA navinuta nebo spirálovitě několikrát velmi specifickým způsobem.
Důležitou roli v tom hrají histony, speciální forma bílkovin. Existuje celkem 5 různých histonů: H1, H2A, H2B, H3 a H4. Dva z posledních čtyř histonů se spojí a vytvoří válcovitou strukturu, oktamer, kolem které se dvojitá šroubovice vinutí asi dvakrát (= super šroubovice). H1 se k této struktuře připojí, aby ji stabilizovala.
Tento komplex DNA, oktameru a H1 se nazývá nukleosom. Několik z těchto nukleosomů je nyní „jako perleťová šňůra“ v relativně krátkých intervalech (10–60 párů bází) za sebou. Úseky mezi chromozomy se nazývají spacerová DNA. Jednotlivé nukleosomy nyní přicházejí znovu do kontaktu prostřednictvím H1, což vytváří další spirálu a tím i kompresi.
Výsledný řetězec je zase ve smyčkách, které jsou stabilizovány páteří vyrobenou z kyselých nehistonových proteinů, také známých jako hertony. Tyto smyčky jsou zase přítomny ve spirálách stabilizovaných proteiny, což vede k poslední fázi komprese. Tento vysoký stupeň komprese se však vyskytuje pouze v kontextu buněčného dělení během mitózy.
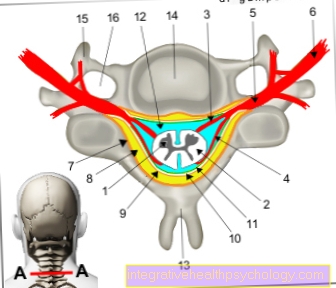
V této fázi můžete také vidět charakteristický tvar chromozomů, který se skládá ze dvou chromatidů. Místo, kde jsou spojeny, se nazývá centroméra. Rozděluje každý metafázový chromozom do dvou krátkých a dvou dlouhých ramen, nazývaných také ramena p a q.
Pokud centromera leží zhruba ve středu chromozomu, nazývá se metacentrický chromozom, pokud leží zcela na jednom z konců akrocentrického chromozomu. Ty mezi nimi se nazývají submetacentrické chromozomy. Tyto rozdíly, které již lze vidět pod světelným mikroskopem, spolu s délkou, umožňují počáteční klasifikaci chromozomů.
Jaké jsou telomery?
Telomery jsou konce chromozomů s opakujícími se sekvencemi (TTAGGG). Ty nenesou žádné relevantní informace, ale slouží spíše jako prevence ztráty relevantnějších částí DNA. S každým buněčným dělením se část chromozomu ztrácí mechanismem replikace DNA.
Telomery jsou tedy v jistém smyslu nárazníkem, který dělí bod, ve kterém buňka ztrácí důležité informace. Pokud jsou telomery buňky kratší než 4 000 párů bází, je zahájena programovaná buněčná smrt (apoptóza). Tím se zabrání šíření vadného genetického materiálu v organismu. Několik buněk má telomerázy, tj. Enzymy, které jsou schopné znovu prodloužit telomery.
Kromě kmenových buněk, ze kterých pocházejí všechny ostatní buňky, jsou to zárodečné buňky a určité buňky imunitního systému. Kromě toho se telomerázy nacházejí také v rakovinných buňkách, a proto se v této souvislosti hovoří o imortalizaci buňky.
Přečtěte si vše o tomto tématu zde: Telomeres - Anatomy, Function & Diseases
Co je to chromatin?
Chromatin označuje celý obsah buněčného jádra, který lze obarvit bází. Kromě DNA tedy termín zahrnuje také určité proteiny, např. Histony a hertony (viz struktura), jakož i určité fragmenty RNA (hn a snRNA).
V závislosti na fázi v buněčném cyklu nebo v závislosti na genetické aktivitě je tento materiál k dispozici v různých hustotách. Hustší forma se nazývá heterochromatin. Pro snazší pochopení by jej tedy bylo možné považovat za „úložnou formu“ a zde opět rozlišovat mezi konstitutivním a fakultativním heterochromatinem.
Konstitutivní heterochromatin je nejhustší forma, která je přítomna ve všech fázích buněčného cyklu v nejvyšší úrovni kondenzace. Tvoří asi 6,5% lidského genomu a nachází se převážně v blízkosti centromer a konců ramen chromozomů (telomery) v malé míře, ale také na jiných místech (hlavně chromozomy 1, 9, 16, 19 a Y) . Kromě toho je většina konstitutivního heterochromatinu umístěna v blízkosti jaderné membrány, tj. Na okrajích buněčného jádra. Prostor uprostřed je vyhrazen pro aktivní chromatin, euchromatin.
Fakultativní heterochromatin je o něco méně hustý a lze jej aktivovat a deaktivovat podle potřeby nebo v závislosti na stadiu vývoje. Dobrým příkladem toho je druhý chromozom X u ženských karyotypů. Jelikož jeden chromozom X je v zásadě dostatečný pro přežití buňky, což je v konečném důsledku dostatečné pro muže, jeden z nich je deaktivován v embryonální fázi. deaktivovaný chromozom X je znám jako Barrovo tělo.
Pouze během buněčného dělení, v kontextu mitózy, zcela kondenzuje, čímž dosahuje nejvyšší komprese v metafázi. Protože se však různé geny čte různě často - koneckonců ne každý protein je vyžadován vždy ve stejném množství - rozlišuje se zde také mezi aktivním a neaktivním euchromatinem.
Přečtěte si více o tomto pod: Chromatin
Haploidní chromozomy
Haploid (řecky haploos = single) znamená, že všechny chromozomy buňky jsou přítomny jednotlivě, tj. Ne v párech (diploidních), jak je tomu obvykle. Jedná se o přirozený stav všech vaječných a spermií, ve kterém se během první meiózy nejprve nerozdělí dva identické chromatidy, nýbrž se nejprve rozdělí všechny páry chromozomů.
Výsledkem je, že po první meióze mají dceřiné buňky u lidí pouze 23 místo obvyklých 46 chromozomů, což odpovídá polovině haploidní sady chromozomů. Protože tyto dceřiné buňky mají stále identickou kopii každého chromozomu sestávajícího ze 2 chromozomů, je nutná druhá meióza, ve které jsou tyto dvě chromatidy od sebe odděleny.
Polytenové chromozomy
Polytenový chromozom je chromozom tvořený velkým počtem geneticky identických chromatidů. Vzhledem k tomu, že tyto chromozomy jsou snadno viditelné i při menším zvětšení, jsou někdy označovány jako obří chromozomy. Předpokladem je endoreplikace, při které se chromozomy uvnitř buněčného jádra několikrát znásobí, aniž by došlo k buněčnému dělení.
Jaké jsou funkce chromozomů?
Chromozom jako organizační jednotka našeho genomu slouží především k zajištění rovnoměrného distribuce zdvojeného genomu mezi dceřinými buňkami během dělení buněk. K tomu je třeba se blíže podívat na mechanismy buněčného dělení nebo buněčného cyklu:
Buňka tráví většinu buněčného cyklu v mezifázi, což znamená celé časové období, ve kterém se buňka hned nerozdělí. To je zase rozděleno na G1, S a G2 fáze.
Fáze G1 (G jako v mezeře, tj. Mezera) bezprostředně následuje po dělení buněk. Zde se buňka opět zvětšuje a plní obecné metabolické funkce.
Odtud může také přejít do fáze G0. To znamená, že se mění na fázi, která již není schopna dělení, a v normálních případech se také velmi mění, aby splnila velmi specifickou funkci (diferenciace buněk). Za účelem splnění těchto úkolů jsou velmi specifické geny čteny intenzivněji, jiné méně nebo vůbec.
Pokud segment DNA není delší dobu potřebný, nachází se často v částech chromozomů, které jsou po dlouhou dobu hustě zabaleny (viz chromatin). Na jedné straně to má za cíl úsporu místa, ale kromě dalších mechanismů genové regulace je to také další ochrana proti náhodnému čtení. Bylo však také pozorováno, že za velmi specifických podmínek mohou diferencované buňky z fáze G0 znovu vstoupit do cyklu.
Po fázi G1 následuje fáze S, tj. Fáze, ve které je syntetizována nová DNA (replikace DNA). Tady musí být celá DNA ve své nejvolnější formě, tj. Všechny chromozomy jsou zcela odvinuty (viz struktura).
Na konci fáze syntézy je celý genetický materiál v buňce přítomen v duplikátu. Protože kopie je stále připojena k původnímu chromozomu přes centromeru (viz struktura), nemluví se o duplikaci chromozomů.
Každý chromozom se nyní skládá ze dvou chromatid namísto jedné, takže během mitózy může později převzít charakteristický tvar X (přesně řečeno, tvar X platí pouze pro metacentrické chromozomy). V následující fázi G2 probíhá okamžitá příprava na buněčné dělení. To zahrnuje také podrobnou kontrolu chyb replikace a přerušení řetězců, které lze v případě potřeby opravit.
V zásadě existují dva typy buněčného dělení: mitóza a meióza. S výjimkou zárodečných buněk všechny buňky organismu vznikají mitózou, jejímž jediným úkolem je tvorba dvou geneticky identických dceřiných buněk.
Meióza má na druhé straně účel generovat geneticky odlišné buňky:
V prvním kroku se rozdělí odpovídající (homologní), ale ne identické chromozomy. Teprve v dalším kroku jsou chromozomy, které se skládají ze dvou identických chromatidů, odděleny a znovu distribuovány do dvou dceřiných buněk, takže nakonec z jedné prekurzorové buňky vzniknou čtyři zárodečné buňky s různým genetickým materiálem.
Forma a struktura chromozomů jsou zásadní pro oba mechanismy: Speciální „proteinová vlákna“, tzv. Vřetenová aparatura, se připojují k vysoce kondenzovaným chromozomům a jemně regulovaným procesem táhnou chromozomy ze střední roviny (rovníkové roviny). na opačné póly buňky kolem jednoho, aby byla zajištěna rovnoměrná distribuce. I malé změny v mikrostruktuře chromozomů zde mohou mít vážné následky.
U všech savců určuje poměr pohlavních chromozomů X a Y také pohlaví potomků. V zásadě vše závisí na tom, zda spermie, které se spojí s vaječnou buňkou, mají chromozom X nebo Y. Vzhledem k tomu, že obě formy spermií jsou vždy produkovány ve přesně stejném rozsahu, je pravděpodobnost vždy vyvážená pro obě pohlaví. Tento náhodný systém zaručuje rovnoměrnější rozdělení pohlaví, než by tomu bylo například v případě faktorů prostředí, jako je teplota.
Zjistěte více o tématu: Dělení buněčných jader
Jak se genetická výbava přenáší přes chromozomy?
Dnes víme, že vlastnosti se dědí prostřednictvím genů, které jsou v buňkách uloženy ve formě DNA. Ty jsou zase rozděleny do 46 chromozomů, na kterých je distribuováno 25 000-30000 lidských genů.
Kromě samotné vlastnosti, která se nazývá fenotyp, existuje také genetický ekvivalent, který se nazývá genotyp. Místo, kde je gen na chromozomu, se nazývá lokus. Protože lidé mají dvojnásobek každého chromozomu, každý gen se také vyskytuje dvakrát. Jedinou výjimkou jsou X-chromozomální geny u mužů, protože Y-chromozom nese pouze zlomek genetické informace nalezené na X-chromozomu.
Různé geny, které jsou na stejném místě, se nazývají alely. Často jsou na jednom místě více než dvě různé alely. Jeden pak mluví o polymorfismu. Taková alela může být jednoduše neškodná varianta (normální varianta), ale také patologické mutace, které mohou být spouštěčem dědičného onemocnění.
Pokud je mutace jediného genu dostatečná ke změně fenotypu, hovoří se o monogenní nebo Mendelově dědičnosti. Mnoho z dědičných znaků se však dědí prostřednictvím několika interagujících genů, a proto je mnohem obtížnější je studovat.
Vzhledem k tomu, že matka i otec předávají dítěti v Mendelianově dědictví jeden ze svých dvou genů, existují v příští generaci vždy čtyři možné kombinace, přičemž tyto mohou být stejné i ve vztahu k jedné vlastnosti. Pokud mají obě alely jedince stejný účinek na fenotyp, je jedinec ve vztahu k této charakteristice homozygotní a charakteristika je odpovídajícím způsobem plně vyjádřena.
Heterozygoti mají dvě různé alely, které mohou vzájemně interagovat různými způsoby: Pokud je jedna alela dominantní nad druhou, zcela potlačuje její expresi a dominantní znak se stává viditelným ve fenotypu. Potlačená alela se nazývá recesivní.
V případě kodominantní dědičnosti se obě alely mohou navzájem neovlivňovat, zatímco v případě přechodné dědičnosti existuje kombinace obou charakteristik. Dobrým příkladem je systém krevních skupin AB0, ve kterém jsou A a B společně dominantní, ale 0 jsou dominantní nad sebou.
Jaká je normální sada chromozomů u lidí?
Lidské buňky mají 22 pohlavně nezávislých párů chromozomů (autozomy) a dva pohlavní chromozomy (gonosomy), takže celkem 46 chromozomů tvoří jednu sadu chromozomů.
Autosomy obvykle přicházejí ve dvojicích. Chromozomy páru mají podobný tvar a sekvenci genů, a proto se označují jako homologní. Dva chromozomy X u žen jsou také homologní, zatímco muži mají chromozom X a Y. Ty se liší formou a počtem přítomných genů takovým způsobem, že již nelze mluvit o homologii.
Zárodečné buňky, tj. Buňky vajíčka a spermií, mají pouze polovinu chromozomů způsobených meiózou, konkrétně 22 jednotlivých autosomů a každý po jednom gonozomu. Vzhledem k tomu, že zárodečné buňky během oplodnění fúzují a někdy vyměňují celé segmenty (křížení), vzniká nová kombinace chromozomů (rekombinace). Všechny chromozomy společně se nazývají karyotyp, který je až na několik výjimek (viz chromozomové aberace) u všech jedinců stejného pohlaví stejný.
Zde se dozvíte vše o daném tématu: Mitóza - jednoduše vysvětleno!
Proč vždy existují páry chromozomů?
V zásadě lze na tuto otázku odpovědět jednou větou: Protože se ukázalo, že je prospěšná.Přítomnost párů chromozomů a princip rekombinace jsou nezbytné pro dědictví z hlediska sexuální reprodukce. Tímto způsobem může zcela nový jedinec náhodou vzniknout z genetického materiálu dvou jedinců.
Tento systém enormně zvyšuje rozmanitost vlastností v rámci druhu a zajišťuje, že se dokáže adaptovat na změněné podmínky prostředí mnohem rychleji a pružněji, než by to bylo možné pouze pomocí mutace a selekce.
Dvojitá sada chromozomů má také ochranný účinek: pokud by mutace genu vedla k selhání funkce, ve druhém chromozomu stále existuje jakási „záložní kopie“. To není vždy dost pro to, aby organismus kompenzoval poruchu, zvláště pokud je mutovaná alela dominantní, ale zvyšuje to její pravděpodobnost. Tímto způsobem se navíc mutace automaticky nepřenáší na všechny potomky, což zase chrání druh před příliš radikálními mutacemi.
Co je to chromozomová mutace?
Genetické vady mohou vznikat z ionizujícího záření (např. Rentgenové záření), chemických látek (např. Benzopyrenu v cigaretovém kouři), určitých virů (např. Virů HP) nebo s nízkou pravděpodobností mohou také vznikat čistě náhodou. Na jeho vývoji se často podílí několik faktorů. V zásadě se takové změny mohou vyskytnout ve všech tělesných tkáních, ale z praktických důvodů je analýza obvykle omezena na lymfocyty (speciální typ imunitní buňky), fibroblasty (buňky pojivové tkáně) a buňky kostní dřeně.
Chromozomová mutace je hlavní strukturální změnou v jednotlivých chromozomech. Nepřítomnost nebo přidání celých chromozomů by na druhé straně byla genomová nebo ploidní mutace, zatímco termín genová mutace označuje poměrně malé změny v genu. Termín chromozomová aberace (latinsky aberrare = odchýlit se) je poněkud širší a zahrnuje všechny změny, které lze detekovat světelným mikroskopem.
Mutace mohou mít velmi odlišné účinky:
- Tiché mutace, tj. Mutace, u nichž změna nemá vliv na jednotlivce nebo jejich potomky, jsou pro chromozomové aberace spíše atypické a častěji se vyskytují v oblasti genových nebo bodových mutací.
- Jeden hovoří o mutaci ztráty funkce, když mutace vyústí ve špatně poskládaný a tedy nefunkční protein nebo vůbec žádný protein.
- Takzvané mutace zesílení funkce mění typ účinku nebo množství produkovaných proteinů tak, že vznikají zcela nové účinky. Na jedné straně je to zásadní mechanismus pro evoluci, a tedy pro přežití druhu nebo pro vznik nových druhů, ale na druhé straně, stejně jako v případě chromosomu Philadelphia, může také rozhodujícím způsobem přispět k vývoj rakovinných buněk.
Nejznámější z různých forem aberací chromozomů jsou pravděpodobně numerické aberace, ve kterých jsou jednotlivé chromozomy přítomny pouze jednou (monosomie) nebo dokonce trojnásobně (trizomie).
Pokud to platí pouze pro jeden chromozom, nazývá se to aneuploidie a celá chromozomová sada je ovlivněna polyploidií (tri- a tetraploidií). Ve většině případů tato nesprávná distribuce vzniká v průběhu vývoje zárodečných buněk nedesepinací (nedisjunkce) chromozomů během dělení buněk (meióza). To vede k nerovnoměrné distribuci chromozomů na dceřiných buňkách a tím k numerické aberaci u dítěte.
Monosomie nepohlavních chromozomů (= autosomů) jsou neslučitelné se životem, a proto se u živých dětí nevyskytují. S výjimkou trizomií 13, 18 a 21 vedou autosomální trizomie téměř vždy k spontánním potratům.
V každém případě, na rozdíl od aberací pohlavních chromozomů, které mohou být také nenápadné, vždy existují závažné klinické příznaky a zpravidla více či méně výrazné vnější abnormality (dysmorfismy).
Taková nesprávná distribuce může nastat také později v životě s mitotickým dělením buněk (všechny buňky kromě zárodečných buněk). Protože kromě postižených buněk existují i nezměněné buňky, hovoří se o somatické mozaice. Pod somatickým (řecky soma = tělo) se rozumí všechny buňky, které nejsou zárodečnými buňkami. Jelikož je ovlivněna pouze malá část buněk těla, příznaky jsou obvykle mnohem mírnější. Proto typy mozaiky často zůstávají po dlouhou dobu nezjištěné.
Zde se dozvíte vše o daném tématu: Mutace chromozomu
Co je chromozomální aberace?
Strukturální chromozomová aberace v zásadě odpovídá definici mutace chromozomu (viz výše). Pokud množství genetického materiálu zůstává stejné a je jednoduše distribuováno odlišně, hovoří se o vyvážené aberaci.
To se často provádí translokací, tj. Přenosem chromozomového segmentu do jiného chromozomu. Pokud se jedná o výměnu mezi dvěma chromozomy, hovoří se o vzájemné translokaci. Protože k produkci proteinů jsou zapotřebí pouze asi 2% genomu, je velmi nízká pravděpodobnost, že takový gen je v bodě zlomu, a tím ztrácí svou funkci nebo je v něm narušen. Proto taková vyvážená aberace často zůstává bez povšimnutí a přenáší se po několik generací.
To však může vést k maldistribuci chromozomů během vývoje zárodečných buněk, což může vést k neplodnosti, spontánním potratům nebo potomkům s nevyváženou aberací.
K nevyvážené aberaci může dojít také spontánně, tj. Bez rodinné anamnézy. Pravděpodobnost, že se dítě narodí živé s nevyváženou aberací, silně závisí na ovlivněných chromozomech a pohybuje se mezi 0 a 60%. To vede ke ztrátě (= deleci) nebo duplikaci (= duplikaci) segmentu chromozomu. V této souvislosti se také hovoří o částečných mono- a trizomiích.
V některých případech k nim dochází společně ve dvou různých oblastech, přičemž pro výskyt klinických příznaků je obvykle rozhodující parciální monosomie. Toto jsou prominentní příklady odstranění Syndrom kočičího výkřiku a syndrom Wolfa-Hirschhorna.
Jeden hovoří o mikrodeleci, když změnu již nelze určit pomocí světelného mikroskopu, tj. Když jde o ztrátu jednoho nebo několika genů. Tento jev je považován za příčinu Prader-Williho syndromu a Angelmanova syndromu a úzce souvisí s vývojem retionoblastomu.
Robertsonova translokace je zvláštní případ:
Dva akrocentrické chromozomy (13, 14, 15, 21, 22) se spojí ve své centromere a po ztrátě krátkých ramen tvoří jediný chromozom (viz struktura). I když to má za následek snížený počet chromozomů, je to označováno jako vyvážená aberace, protože ztrátu krátkých ramen v těchto chromozomech lze snadno kompenzovat. I zde jsou účinky často patrné až v následujících generacích, protože existuje velmi vysoká pravděpodobnost potratů nebo živých dětí s trizomií.
Pokud v chromozomu dojde ke dvěma zlomům, může se stát, že se mezilehlý segment otočí o 180 ° a začlení se do chromozomu. Tento proces, známý jako inverze, je nevyvážený, pouze pokud bod zlomu leží v aktivním genu (2% z celkového genetického materiálu). V závislosti na tom, zda je centromera uvnitř nebo vně obráceného segmentu, jde o peri- nebo paracentrickou inverzi. Tyto změny mohou také přispět k nerovnoměrnému rozložení genetického materiálu na zárodečné buňky.
V paracentrické inverzi, ve které centromera není v obráceném segmentu, se mohou objevit také zárodečné buňky se dvěma nebo bez centromery. Výsledkem je, že odpovídající chromozom je ztracen během prvních buněčných dělení, což téměř jistě vede k potratu.
Vložení je instalace fragmentu chromozomu na jiné místo. I zde jsou potomci primárně postiženi podobným způsobem. Kroužkový chromozom může nastat zejména po odstranění koncových částí. Typ a velikost sekvencí jsou rozhodující pro závažnost příznaků. Navíc to může vést k nesprávnému rozdělení a tím zase k mozaikovým typům v tělních buňkách.
Pokud se chromafom metafáze během buněčného dělení nesprávně oddělí, mohou vzniknout izochromozomy. Jedná se o dva přesně stejné chromozomy, které se skládají pouze z dlouhých nebo pouze krátkých ramen. V případě chromozomu X se to může projevit jako Ulrich-Turnerův syndrom (monosomie X).
Přečtěte si více informací o tomto tématu: Chromozomální aberace
Trizomie 21
Trizomie 21, lépe známá jako Downův syndrom, je pravděpodobně nejčastější numerickou chromozomální aberací u živě narozených dětí, přičemž muži jsou postiženi o něco častěji (1,3: 1).
Pravděpodobnost výskytu trizomie 21 závisí na různých demografických faktorech, jako je průměrný věk při narození matek, a v jednotlivých regionech se mírně liší.
95% trizomie 21 vzniká v důsledku chyby dělení v kontextu meiózy (dělení zárodečných buněk), konkrétně nondisjunkce, tj. Selhání oddělení sesterských chromatidů.
Tito jsou známí jako volné trizomie a vznikají 90% u matky, 5% u otce a dalších 5% v embryonálním genomu.
Další 3% jsou důsledkem nevyvážených translokací buď na chromozomu 14 nebo jako 21; Translokace 21, vytvoření normálního a dvojitého chromozomu 21. Zbývající 2% tvoří mozaikové typy, u nichž trizomie nevznikla v zárodečných buňkách, a proto neovlivňuje všechny buňky těla. Typy mozaik jsou často tak mírné, že mohou zůstat zcela nezjištěné po dlouhou dobu.
V každém případě by mělo být provedeno vyšetření chromozomu, aby bylo možné odlišit symptomaticky identickou volnou trizomii od případně zděděné translokační trizomie. Potom může následovat rodinná historie předchozích generací.
Zajímá vás toto téma? Přečtěte si o tom následující článek: Trizomie 21
Trizomie 13
Trizomie 13 nebo Patauův syndrom má frekvenci 1: 5 000 a je mnohem vzácnější než Downov syndrom. Příčiny (volné trizomie, translokace a typy mozaik) a jejich procentuální rozdělení jsou do značné míry totožné.
Teoreticky lze téměř všechny případy diagnostikovat prenatálně pomocí ultrazvuku nebo testu PAPP-A. Jelikož test PAPP-A nemusí být nutně součástí rutinních vyšetření, přibližně 80% případů ve střední Evropě je diagnostikováno před narozením.
Na ultrazvuku je již vidět růstový zbytek, bilaterální rozštěp rtu a patra a neobvykle malé oči (mikroftalmie). Kromě toho jsou obvykle přítomny malformace předního mozku a obličeje různého stupně závažnosti (holoprosencephaly).
Zatímco v lobární formě jsou mozkové hemisféry téměř úplně oddělené a jsou vytvořeny laterální komory, v semi-lobární formě je často oddělena pouze zadní část mozku a postranní komory chybí. V nejtěžší formě, alobární formě, nedochází k oddělení mozkových hemisfér.
Kojenci s polo- nebo alobárním tvarem obvykle umírají ihned po narození. Po jednom měsíci se úmrtnost pohybuje kolem 50% živě narozených. Až do věku 5 let se úmrtnost na trizomii 13 zvyšuje na 90%. Kvůli malformacím v mozku zůstávají ve většině případů nemocní lidé na celý život upoutáni na lůžko a neumí mluvit, a proto jsou závislí na plné péči. Kromě toho mohou také existovat dalekosáhlé fyzické projevy Trismoie 13.
Přečtěte si více o tomto tématu na: Trizomie 13 u nenarozeného dítěte
Trizomie 16
V zásadě je trizomie 16 nejběžnější trizomií (přibližně 32% všech trizomií), ale žijící děti s trizomií 16 jsou velmi vzácné. Obecně platí, že živě narozené děti se vyskytují pouze v částečných trizomiích nebo mozaikových typech. Mezi trizomiemi je však nejčastěji zodpovědná za mrtvě narozené děti: 32 ze 100 potratů v důsledku chromozomálních aberací lze vysledovat zpět k této formě trizomie.
Proto byly dokumentovány hlavně prenatální, tj. Prenatální, identifikovatelné vlastnosti. Za zmínku stojí různé srdeční vady, zpomalený růst, jediná pupečníková tepna (jinak zdvojnásobená) a zvýšená průhlednost krku, což je vysvětleno hromaděním tekutin v důsledku dosud nevyvinutého lymfatického systému a zvýšené pružnosti kůže v této oblasti. Kromě toho fyziologická pupeční kýla, tj. Dočasné přemístění velké části střeva přes pupek ven, často neprobíhá správně, což se označuje jako zlomení omfalokély nebo pupečníku.
Ohybová kontraktura se zkříženými prsty může být také často detekována na ultrazvuku. U několika živě narozených dětí je patrná generalizovaná svalová hypotenze, tj. Celková svalová slabost. To vede k slabosti při pití a může zajistit, že kojenec musí být krmen uměle. Často se také vyskytuje čtyřprstá brázda, která je tak charakteristická pro trizomie. I zde frekvence výskytu trizomie přímo souvisí s věkem matky.
Trizomie 18
Edwardsův syndrom, tj. Trizomie 18, se vyskytuje s frekvencí 1: 3000. U prenatální diagnostiky je to stejné jako u Patauova syndromu: i zde by stejná vyšetření umožnila úplné nalezení všech pacientů před narozením. Příčiny a jejich rozdělení lze srovnávat s jinými trizomiemi (viz trizomie 21).
Kromě toho se u trizomie 18 vyskytují částečné trizomie, které stejně jako mozaikové typy vedou k mnohem mírnějším klinickým průběhům. Související dysmorfismy jsou také extrémně charakteristické pro Edwardsův syndrom: Při narození mají pacienti výrazně sníženou tělesnou hmotnost o 2 kg (normální: 2,8–4,2 kg), ustupující široké čelo, obecně nedostatečně vyvinutá dolní polovina obličeje s malými ústy otevírání, úzké štěrbiny očních víček a otočení dozadu, tvarově změněné uši (faunovo ucho). Znatelná je také zadní část hlavy, která je u novorozence neobvykle silně vyvinutá. Žebra jsou neobvykle úzká a křehká. Novorozenci mají také trvalé napětí (tón) celého svalstva, které však u přeživších po prvních týdnech ustupuje.
Dalším charakteristickým rysem je zkřížení 2. a 5. prstu přes 3. a 4. s celkovým počtem zatlučených prstů, zatímco chodidla jsou neobvykle dlouhá (uplynulá), mají obzvláště výraznou patu, zakrnělé nehty a zadní palec. .
Závažné orgánové malformace jsou časté a obvykle se vyskytují v kombinaci: srdeční a ledvinové vady, nesprávné nasazení (malrotace) střeva, adheze peritonea (mesenterium commune), okluze jícnu (atrézie jícnu) a mnoho dalších.
Z důvodu těchto malformací se úmrtnost pohybuje kolem 50% během prvních 4 dnů, pouze asi 5–10% se dožívá věku přes rok. Absolutní výjimkou je přežití do dospělosti. V každém případě je mentální postižení velmi výrazné a neumí mluvit, je upoutáno na lůžko a je inkontinentní, takže je zcela závislé na pomoci zvenčí.
Podrobnější informace o trizomii 18 najdete také v našem podrobném článku na toto téma:
- Trizomie 18 (Edwardsův syndrom)
- Trizomie 18 u nenarozeného dítěte
Trizomie X
Trizomie X je nejnápadnější formou numerické chromozomální aberace, vzhled postižených, které jsou logicky všechny ženy, se od ostatních žen příliš neliší. Některé vynikají tím, že jsou obzvláště vysoké a mají trochu „kypré“ rysy obličeje. Mentální vývoj může být také do značné míry normální, od hraničního normálu po mírné mentální postižení.
Tento deficit inteligence je však o něco závažnější než u ostatních trizomií pohlavních chromozomů (XXY a XYY). S frekvencí 1: 1000 to ve skutečnosti není tak vzácné, ale protože trizomie obvykle není spojena s klinicky významnými příznaky, většina žen s tímto onemocněním pravděpodobně nikdy nebude diagnostikována po celý svůj život.
Nosiče jsou většinou objeveny náhodou během rodinné kontroly nebo během prenatální diagnostiky.Plodnost lze mírně snížit a míru aberací pohlavních chromozomů v příští generaci lze mírně zvýšit, takže pokud chcete mít děti, doporučuje se genetické poradenství.
Stejně jako u ostatních trizomií se trizomie X nejčastěji vyvíjí jako volná trizomie, tj. Kvůli nedostatečnému dělení (nondisjunkce) sesterských chromatid. I zde obvykle vzniká během zrání mateřských vaječných buněk, i když pravděpodobnost s věkem roste.
Syndrom křehkého X
U mužů je preferován syndrom křehkého X nebo Martin Bell, protože mají pouze jeden chromozom X, a proto jsou touto změnou více ovlivněni.
Vyskytuje se s frekvencí 1: 1250 u živě narozených mužů v jednom roce, což z něj činí nejběžnější formu nespecifické mentální retardace, tj. Všechny mentální handicapy, které nelze popsat zvláštním syndromem s typickými znaky.
Syndrom křehkého X se obvykle může objevit u dívek v poněkud slabší formě, což je způsobeno náhodnou inaktivací jednoho z X chromozomů. Čím vyšší je podíl vypnutého zdravého chromozomu X, tím silnější jsou příznaky.
Většinu času jsou však ženy nositelkami premutace, která dosud nevyvolává žádné klinické příznaky, ale výrazně zvyšuje pravděpodobnost úplné mutace u jejich synů. Ve velmi vzácných případech mohou být muži také nositeli premutace, kterou pak mohou předat pouze dcerám, které jsou však obvykle klinicky zdravé (Shermanův paradox).
Tento syndrom je vyvolán extrémně zvýšeným počtem tripletů CGG (určitá základní sekvence) v genu FMR (fragile-site-mental-retardation); namísto 10–50 kopií premutace 50–200, když je plně vyvinuta 200– 2000 výtisků.
Pod světelným mikroskopem to vypadá jako zlomenina dlouhé paže, což dalo tomuto syndromu jeho jméno. To vede k deaktivaci postiženého genu, což zase způsobuje příznaky.
Postižení lidé vykazují zpomalený vývoj řeči a pohybu a mohou vykazovat problémy v chování, které mohou jít směrem k hyperaktivitě, ale také autismu. Čistě vnější abnormality (známky dysmorfismu) jsou dlouhá tvář s výraznou bradou a vyčnívajícími ušima. S pubertou jsou varlata často značně zvětšena (makroorchidie) a rysy obličeje se stávají hrubšími. Existuje mírná akumulace psychologických abnormalit a zvláště časná menopauza mezi ženskými nositelkami premutace.
Co je to analýza chromozomu?
Analýza chromozomů je proces v cytogenetice, pomocí kterého lze detekovat numerické nebo strukturní aberace chromozomů.
Taková analýza by se použila například v případě bezprostředního podezření na chromozomální syndrom, tj. V případě malformací (dysmorfismus) nebo mentálního postižení (retardace), ale také v případě neplodnosti, pravidelných potratů (potratů) a také některé druhy rakoviny (např. lymfomy nebo leukémie).
To obvykle vyžaduje lymfocyty, speciální typ imunitní buňky, který se získává z krve pacienta. Protože tímto způsobem lze získat pouze relativně malé množství, jsou buňky stimulovány k dělení fytohemaglutininem a lymfocyty mohou být poté kultivovány v laboratoři.
V některých případech se místo toho podobným způsobem odebírají vzorky (biopsie) z kůže nebo míchy. Cílem je získat co nejvíce materiálu DNA, který je v současné době uprostřed buněčného dělení. V metafázi jsou všechny chromozomy uspořádány v jedné úrovni přibližně ve středu buňky, aby byly v dalším kroku, anafáze, přitahovány na opačné strany (póly) buňky.
V tomto okamžiku jsou chromozomy obzvláště těsně zabalené (vysoce kondenzované). Přidá se vřetenový jed kolchicin, který funguje přesně v této fázi buněčného cyklu, takže se hromadí metafázové chromozomy. Poté se izolují a obarví pomocí speciálních metod barvení.
Nejběžnější je GTG pruhování, při kterém jsou chromozomy ošetřeny trypsinem, trávicím enzymem a pigmentem Giemsa. Obzvláště hustě zbarvené oblasti a oblasti bohaté na adenin a tymin jsou zobrazeny tmavě.
Výsledné G-pásy jsou charakteristické pro každý chromozom a ve zjednodušeném smyslu jsou považovány za oblasti s menším počtem genů. Obrázek takto obarvených chromozomů se pořídí v tisíckrát zvětšeném stavu a pomocí počítačového programu se vytvoří karyogram. Kromě pásového vzoru se používá velikost chromozomu a poloha centromery, které pomáhají odpovídajícím způsobem uspořádat chromozomy. Existují však i jiné způsoby páskování, které mohou mít velmi odlišné výhody.
Doporučení redakčního týmu
Další obecné informace najdete v následujících článcích:
- Dělení buněčných jader
- Funkce buněčného jádra
- Trizomie 21
- Genetické nemoci











.jpg)